| Question | Short Answer |
|---|---|
| What is the first step for cam photosynthesis: why succulents open stomata at night? | Start by matching the symptom to the plant, substrate, light, and season before changing watering or treatment. |
| What should be avoided? | Avoid changing several variables at once; correct the limiting factor and observe the plant before escalating. |
| Which care factor matters most? | Match the plant to its light, substrate, pot size, and season. |
| When should the plant be checked again? | Recheck after one to two weeks unless tissue is actively collapsing. |
Quick Answer
The short answer: CAM plants open their stomata at night (not day) to fix CO2, store it as malic acid, then use it during the day with stomata closed. This uses 3-6x less water than normal photosynthesis.
Best first step: Water in the evening or early morning, not midday - that's when stomata are open and the plant can actually absorb water.
Avoid: Keeping plants in air-conditioned rooms below 20% humidity - CAM stomata won't open in dry air, and carbon gain drops to zero.
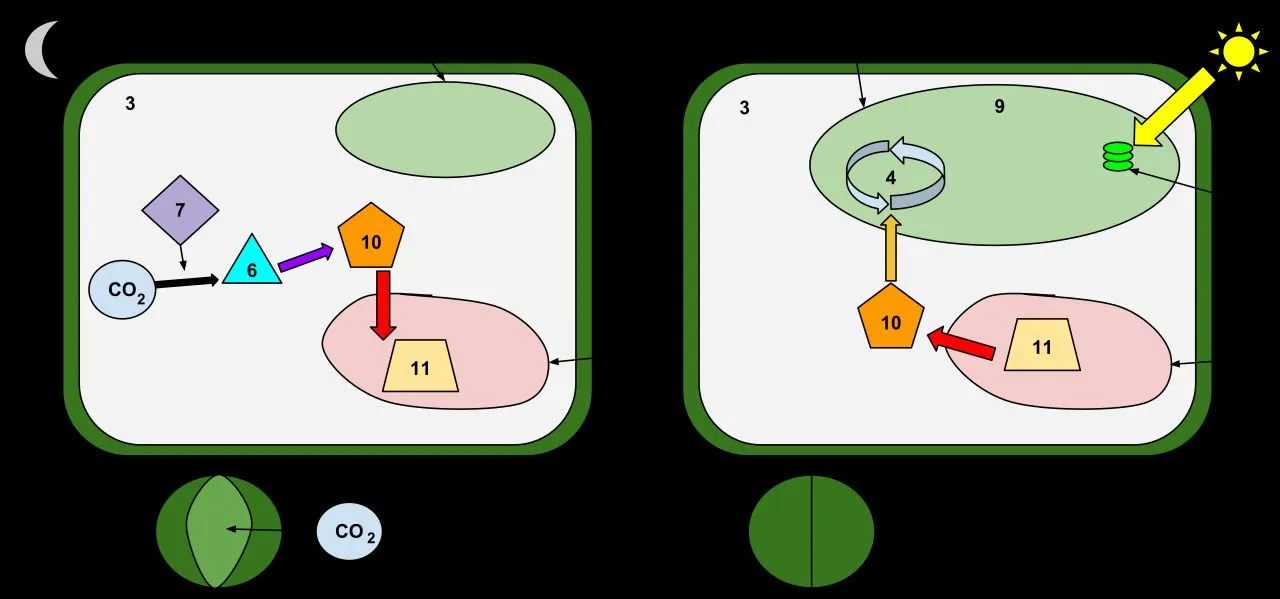
Most temperate plants open their stomata in the morning sun and close them at dusk. Succulents do the opposite. They keep stomata sealed through the heat of the day, then open them in the dark, fix carbon dioxide overnight, store the carbon as malic acid in the vacuole, and run the rest of photosynthesis the next day with the stomata closed. The pathway is called Crassulacean Acid Metabolism, or CAM, and it is the single largest reason succulents survive on a fraction of the water that other plants need. Here is the rest of the picture.
I'm Dr. Elena Martín, a Certified Advanced Cactus & Succulent Horticulturist and former curator of the succulent collection at the Jardín Botánico de Córdoba. The cultivation mistakes I see most often are usually a mismatch between an indoor environment and the night-time biochemistry of a CAM plant. Once you understand what is happening between dusk and dawn, the watering and temperature decisions become obvious.
Part of the Beginner's Guide to Succulents.
What CAM is biologically
Crassulacean Acid Metabolism is named after the Crassulaceae, the family in which it was first characterised in the early twentieth century. It is a temporal separation of two photosynthetic steps that other plants run in parallel during daylight: gas exchange and carbon fixation by Rubisco.
In a typical C3 plant, leaves open their stomata in daylight, take in atmospheric CO2, and feed it directly into the Calvin cycle, where Rubisco fixes it onto a five-carbon sugar precursor. CO2 entry and Rubisco fixation happen at the same time, in the same cell, in the same daylight hours. The cost is water: a leaf with daylight stomata loses several hundred grams of water vapour for every gram of CO2 it absorbs.
CAM plants moved the gas exchange step to night. Their stomata open from dusk to dawn, when humidity is higher and air temperature is cooler, so transpirative water loss per unit CO2 absorbed is far smaller. The CO2 that enters at night is not handed straight to Rubisco. Instead, an enzyme called PEP carboxylase catches it onto a three-carbon acceptor, phosphoenolpyruvate, producing oxaloacetate. Oxaloacetate is then reduced to malate, which is shunted into the cell vacuole as malic acid and accumulates there overnight.
The chemistry is literal. The leaf becomes acidic by morning. Vacuolar pH typically falls from about 6.5 in the late afternoon to about 4.0 by sunrise, a thousand-fold change in proton concentration. Nineteenth-century botanists noticed this directly by tasting Bryophyllum and Kalanchoe leaves at different hours, well before any of the biochemistry was known.
When the sun rises, the cycle reverses. Stomata close to conserve water. The malic acid stored in the vacuole moves back into the cytoplasm, is decarboxylated, and releases CO2 inside the leaf. Rubisco, sitting in the chloroplast stroma, sees a high internal CO2 concentration and runs the Calvin cycle on this internally supplied carbon, all behind closed stomata.
The 24-hour cycle in detail
The CAM cycle is conventionally divided into four phases over the day-night cycle.
Phase I runs through the night. Stomata are open. PEP carboxylase fixes atmospheric CO2 into oxaloacetate, then malate, which fills the vacuole. Leaf pH falls steadily. This is the period during which the plant is genuinely taking gas from outside, and it is the only window in which environmental humidity, temperature, and ventilation directly affect carbon gain.
Phase II is the brief transition at dawn. Stomata may stay open for an hour or two while light is weak and atmospheric CO2 can still feed Rubisco directly. Internal CO2 begins to rise as malate decarboxylation starts.
Phase III covers the bulk of the daytime. Stomata are closed. The vacuole pumps malic acid back out into the cytoplasm. Decarboxylation, by NADP-malic enzyme or PEP carboxykinase depending on the species, releases CO2 inside the leaf at concentrations several times higher than ambient air. Rubisco fixes this internal CO2 into the Calvin cycle. Leaf pH rises through the morning and stabilises near neutral by mid-afternoon.
Phase IV is the late-afternoon transition. The vacuolar malate store is exhausted, internal CO2 falls, and the plant may briefly reopen stomata to take a small amount of atmospheric CO2 directly before night begins.
This four-phase rhythm runs on a circadian clock. In laboratory conditions, stomatal aperture continues cycling on its 24-hour pattern even under continuous darkness or continuous light.
How it differs from C3 and C4
Three photosynthetic strategies dominate land plants, and the differences matter for anyone keeping CAM plants alongside houseplants from other families.
C3 photosynthesis is the ancestral pathway used by most temperate plants, including grasses, broadleaf trees, and most vegetable crops. CO2 enters open daytime stomata and is fixed directly by Rubisco onto a five-carbon sugar to produce two molecules of a three-carbon compound (hence the name). Water-use efficiency is poor, on the order of 700 grams of water lost per gram of carbon fixed.
C4 photosynthesis is found in tropical grasses (maize, sugarcane, sorghum) and a scattering of dicots. C4 plants also use PEP carboxylase to make a four-carbon acid, but they separate the steps in space rather than time: mesophyll cells fix CO2 into malate, then pass it to bundle-sheath cells where Rubisco runs at high internal CO2. Water-use efficiency is moderate, around 300 g of water per gram of carbon, and the maximum rate per unit leaf area is the highest of the three pathways.
CAM is the third route, and it pays the largest water-saving premium. Water-use efficiency typically runs three to six times higher than C3 plants, with values around 25 grams of water lost per gram of carbon fixed in well-adapted species, against C3's 700 g/g. The trade-off is that the maximum photosynthetic rate per unit leaf area is much slower than either C3 or C4. Rubisco only sees CO2 during the day, when supply depends on the size of the malate store built up overnight, and that supply is limited. CAM plants are therefore slow growers per unit leaf area, but they hold positive carbon balance under conditions that would kill a C3 plant by drought.
Why this matters for cultivation
The first practical consequence is timing. A CAM plant measured at the wrong moment is a different plant from itself measured at the right moment. Leaf turgor, stomatal aperture, internal CO2, and respiration rates all swing through the cycle. If you check a Mammillaria at noon and the body feels slightly soft, you are reading a stomata-closed, malate-depleting state. The same plant at 6 a.m., with vacuoles at peak acidity, feels firmer. Diagnostic squeeze tests work better at dawn than at midday.
The second is night temperature. Daytime warmth matters less for CAM photosynthesis than the temperature during the night-time gas-exchange window. PEP carboxylase activity and stomatal opening peak between roughly 12 and 18 °C at night for most temperate-cultivated species. Below about 5 °C, enzyme activity collapses and stomata may not open at all. Above about 25 °C, transpirative loss climbs even at night, and the water-saving advantage erodes. A heated room held at 22 °C around the clock can give worse photosynthetic performance than a cooler space that drops to 15 °C overnight, even with similar total light.
The third is night humidity. CAM stomata close in dry air regardless of temperature, which protects the plant from running its night-time uptake at a loss. Desert habitat humidity often sits between 30 and 50 percent at night, and CAM plants gas-exchange comfortably under those conditions. An air-conditioned indoor room can drop to 20 percent relative humidity overnight, especially in winter with central heating running. Under that air, stomata may not open even when temperature and light cues are correct, and carbon gain falls toward zero. A hygrometer at bench height is more useful than a thermometer for diagnosing slow growth.
The fourth concerns watering rhythm, and it links directly to the wet-dry rhythm covered in the Beginner's Guide to Succulents. CAM plants tolerate long dry intervals because the pathway is built for them. Watering more often than the substrate dries does not generally damage the plant, but it wastes water that the plant cannot use, and in cool conditions it creates the saturated, oxygen-poor root zone that opportunistic Pythium prefers. Water on the substrate's clock, not on a calendar.
Plant families that use CAM
CAM is not rare. It has evolved independently many times across at least 35 plant families and roughly 6 percent of vascular plant species. Among the families relevant to succulent collectors, the major CAM users are:
- Crassulaceae: the family that gave the pathway its name. Effectively all Crassula, Sedum, Sempervivum, Echeveria, Aeonium, Kalanchoe, Graptopetalum, and Pachyphytum species use CAM, though some show facultative behaviour (see below).
- Cactaceae: virtually all true cacti, from the small Mammillaria and Rebutia to the columnar Carnegiea gigantea of the Sonoran Desert and the epiphytic Schlumbergera and Rhipsalis.
- Aizoaceae: all members of the mesemb family, including Lithops, Conophytum, Pleiospilos, Faucaria, and the broader Mesembryanthemum group.
- Agavaceae: most Agave species, plus relatives like Yucca and Hesperaloe. Pineapple (Ananas comosus) is also a CAM crop of commercial scale.
- Asphodelaceae: many Aloe, Haworthia, and Gasteria run CAM, although some show C3-like behaviour under well-watered greenhouse conditions.
- Euphorbiaceae: the succulent African Euphorbia lineage.
- Apocynaceae: succulent stapeliads such as Stapelia, Huernia, Hoya, and Ceropegia.
Outside the succulents, CAM also appears in many epiphytic orchids and in Tillandsia bromeliads.
CAM-idling: facultative CAM
CAM is not always running flat out. Plants can shift along a continuum from full CAM to partial CAM to a state called CAM-idling, and some species can switch the pathway on and off in response to water status.
Under severe drought, many CAM plants enter CAM-idling. Stomata stay closed at night as well as during the day. The plant continues to photosynthesise, but only on CO2 released internally by its own respiration, which it recaptures and refixes through the malate cycle. No new carbon enters the leaf. The plant survives without gaining mass and without losing water vapour beyond cuticular transpiration. This is the state in which a Sempervivum under July sun on a south-facing wall, or a Mammillaria in a Sonoran summer, can sit for weeks waiting for rain. Idling is not dormancy; it is active maintenance with the gas-exchange tap shut off entirely.
Some species are obligate CAM, meaning they run the full nocturnal pathway throughout life. Most cacti and most Aizoaceae fall into this group. Other species are facultative, switching between C3 photosynthesis when water is available and CAM when it is not. The textbook example is Mesembryanthemum crystallinum, a coastal annual ice plant studied since the 1970s as a model for the C3-to-CAM transition. Well-watered, in mild conditions, M. crystallinum photosynthesises as a C3 plant. Subjected to drought or high salinity, it induces CAM enzymes within days and switches the entire pathway to nocturnal gas exchange.
Several Sedum species show similar plasticity. Sedum album, Sedum acre, and the broader European stonecrop group can run partial or full CAM under drought stress, then return to mostly C3 behaviour with regular rain. The implication for cultivation is that a stressed facultative plant may not look stressed in the way a houseplant does. It is conserving water and protecting itself, not failing. Returning it to comfortable conditions usually returns it to C3 behaviour without intervention.
The CAM strategy is best understood as a set of dials rather than a single switch. A grower who mistakes a healthy CAM-idling plant for a thirsty one, and waters into the closed-stomata state, can shift the substrate from helpful to dangerous in a single afternoon.
See also
- A Beginner's Guide to Succulents: the broader foundation that makes the CAM cycle visible in everyday cultivation
- The Wet-Dry Cycle Explained: how the night-time gas exchange of a CAM plant interacts with the rhythm of substrate moisture
- Winter Watering Protocol: why short photoperiods and cool nights collapse CAM activity and reduce water demand to nearly zero
FAQ
Why should I water succulents at night?
CAM stomata open at night, so the plant can actually absorb water. Watering at midday (when stomata are closed) wastes water and sits in the pot.
Does night temperature affect my succulent?
Yes. PEP carboxylase (the night-time CO2-fixing enzyme) works best at 12-18°C. Below 5°C it stops working; above 25°C water loss increases even at night.
What happens if my room is too dry?
Dry air (<30% humidity) causes CAM stomata to stay closed even at night. The plant can't take up CO2 or water. Use a hygrometer to check - low humidity is why some indoor succulents struggle.
Can succulents switch off CAM photosynthesis?
Some facultative CAM plants (like Sedum) can switch to normal C3 photosynthesis when well-watered, then switch back to CAM during drought. This is why they look fine even in dry conditions.
Frequently Asked Questions
What is the first step for cam photosynthesis: why succulents open stomata at night?
Start by matching the symptom to the plant, substrate, light, and season before changing watering or treatment.
What should be avoided?
Avoid changing several variables at once; correct the limiting factor and observe the plant before escalating.
Which care factor matters most?
Match the plant to its light, substrate, pot size, and season. Most succulent failures trace to a mismatch between drying speed and the plant's current growth rate.
When should the plant be checked again?
Recheck after one to two weeks unless tissue is actively collapsing. Stable firmness and new growth are better signs than a fixed calendar interval.